Mitochondrial diseases: from molecular mechanisms to therapeutic advances

线粒体对细胞功能和活力至关重要,是新陈代谢和信号传递的中心枢纽。它们拥有各种新陈代谢和质量控制机制,对维持正常的细胞活动至关重要。线粒体或核 DNA(编码线粒体蛋白或其他内容物)的各种突变都可能导致线粒体遗传疾病。这些遗传缺陷可能导致线粒体功能和代谢的崩溃,例如氧化磷酸化的崩溃,这是线粒体最关键的功能之一。线粒体疾病是一类常见的遗传性疾病,具有明显的表型和遗传异质性。临床症状可表现在全身各个系统和器官,严重程度和形式各不相同。线粒体与线粒体疾病之间的关系错综复杂,导致人们对这些疾病的基因型与表型之间的相关性认识不足,从而在历史上给诊断和治疗带来了挑战,并常常导致不尽人意的临床结果。然而,近期研究和技术的进步极大地改善了我们对这些疾病的理解和管理。线粒体相关疗法的临床转化正在积极推进。本综述重点介绍线粒体的生理机制、线粒体疾病的发病机制以及潜在的诊断和治疗应用。此外,本综述还讨论了线粒体遗传疾病的未来前景。
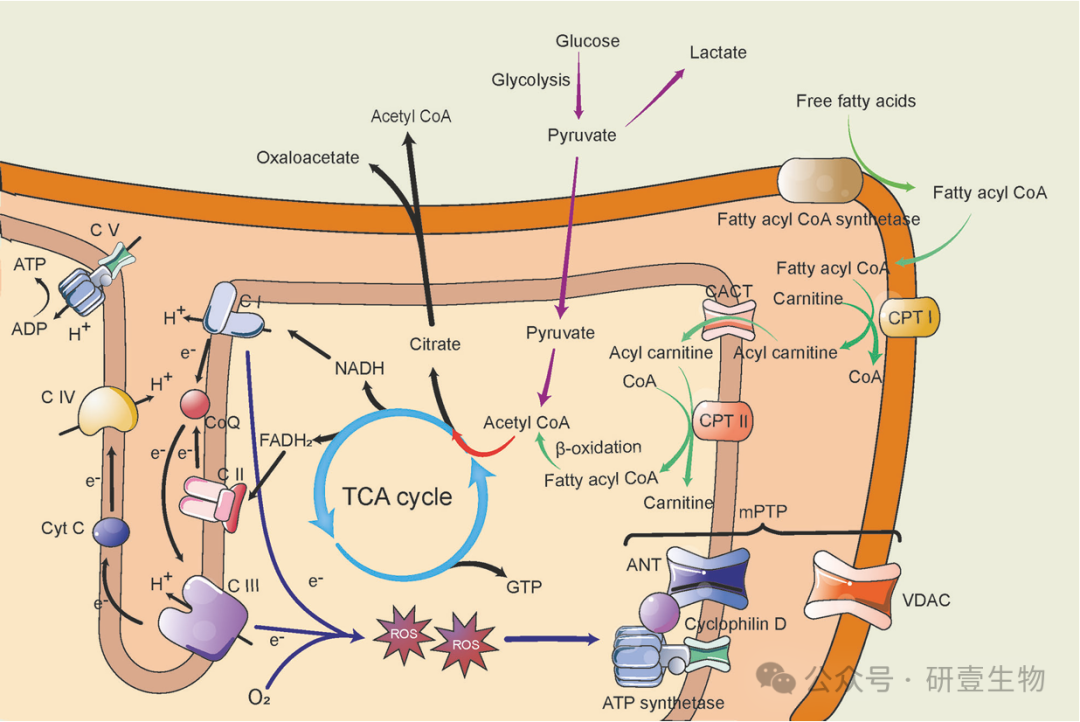
作为生物能的中心枢纽,线粒体利用 TCA 循环产生的 NADH 和 FADH2,通过电子传递和跨呼吸链复合物的 H+ 梯度产生 ATP。复合体 I 和 III 是 mtROS 的主要来源,可导致氧化损伤或信号转导。mtROS 还能诱导 mPTP 开放。葡萄糖和脂质(通过 β-氧化)都有助于 TCA 循环。柠檬酸盐可穿过线粒体膜,使乙酰-CoA 被运输到细胞质中,以发挥各种功能。
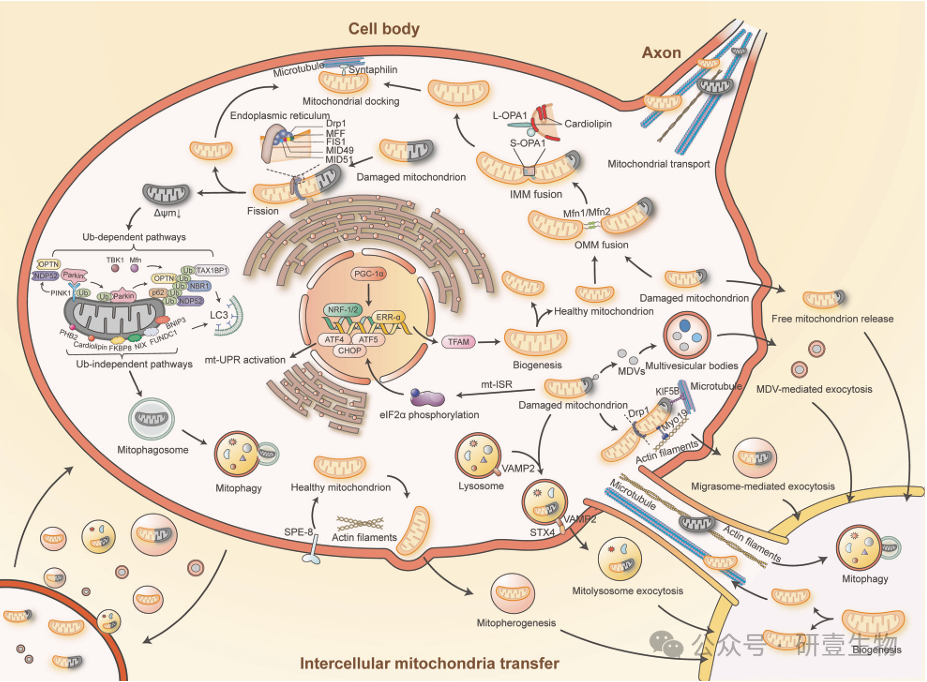
线粒体利用细胞间和细胞内的质量控制机制来维持平衡和氧化还原平衡。这些机制包括线粒体生物生成、融合、裂变、轴突运输、对接、有丝分裂、线粒体综合应激反应和细胞间线粒体转移。
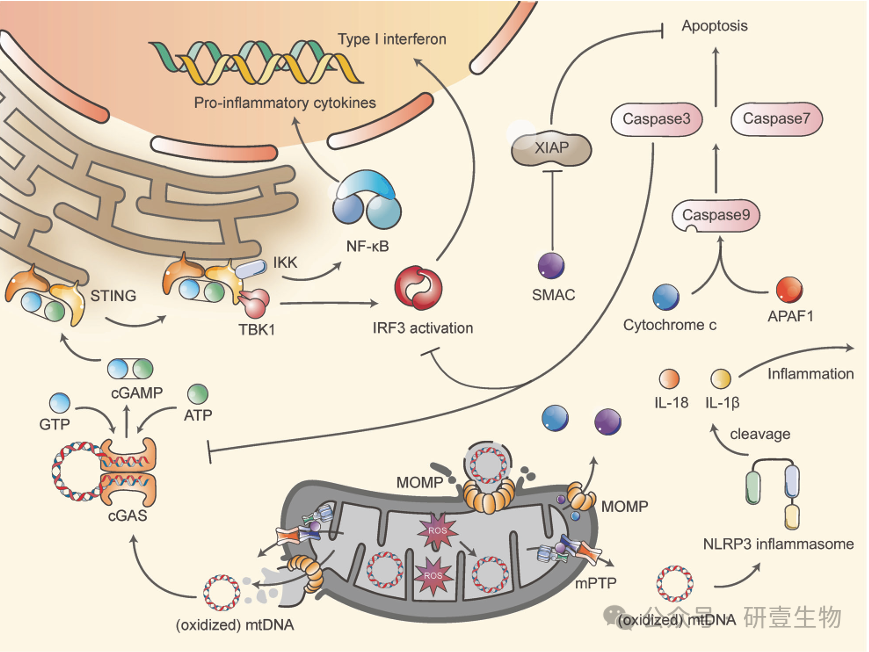
如果 IMS 或基质中的成分泄漏到细胞质中,就会引发细胞凋亡或炎症,这主要是由于 线粒体膜通透性转换孔 (mPTP )和线粒体外膜透化( MOMP) 的作用。MOMP 形成和 mPTP 打开后,细胞色素 C、SMAC 和 mtDNA 会释放到细胞质中。细胞色素 C 与 APAF1 相互作用,激活 caspase 9,启动 caspase 级联反应,导致细胞凋亡。SMAC 通过抑制 XIAP 加速这一过程。cGAS 酶与 mtDNA 结合后,由 ATP 和 GTP 生成 cGAMP,激活 cGAS-STING 信号通路,诱导 I 型干扰素表达和 NF-κB 激活。NLRP3 炎性体还能与(氧化的)mtDNA 结合,促进 IL-1β 和 IL-18 的裂解。然而,在细胞凋亡过程中,caspase 3 会裂解 cGAS 和 IRF3,从而抑制炎症。
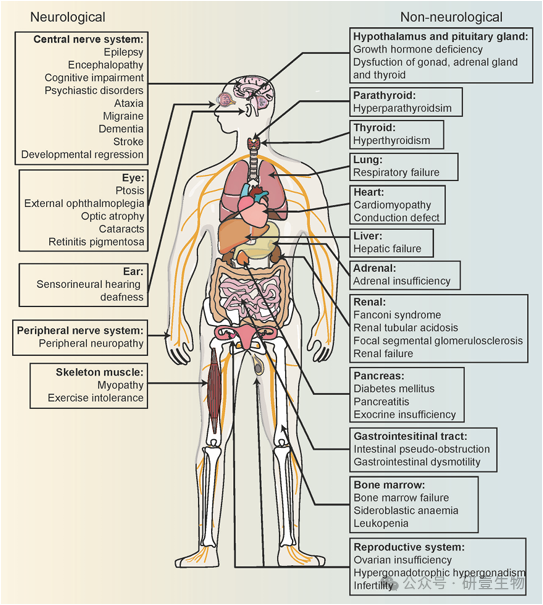
由于线粒体无处不在,线粒体疾病可能出现在人体的任何组织中。骨骼肌和大脑等能量需求较高的组织和器官尤其容易受到氧化磷酸化缺陷的影响,从而导致线粒体疾病中常见的肌病和脑病等表现。与这些疾病相关的症状多样且多变,增加了误诊的风险。
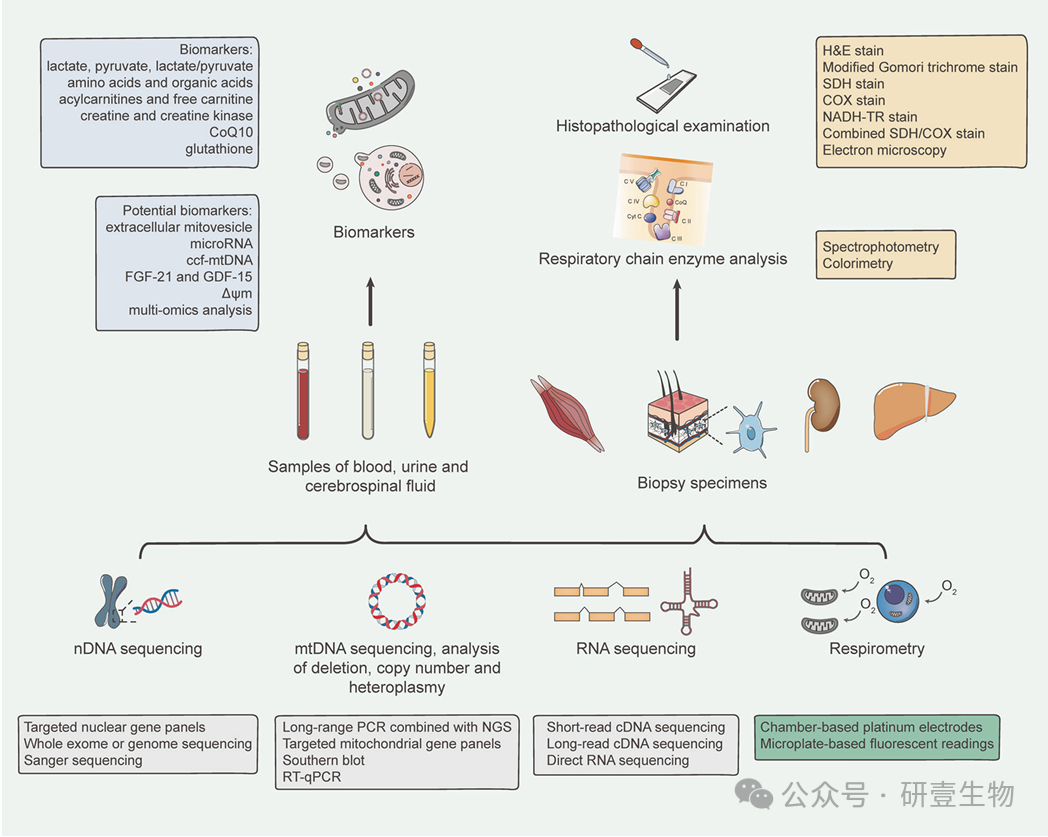
诊断策略已从活检优先发展到基因优先。初步筛查应使用血液、尿液和脑脊液样本。生物标志物检测在这一过程中至关重要。对于疑似线粒体疾病,mtDNA 测序和分析是首选方法,而对于 mtDNA 多缺失、缺失或早期症状的病例,则应考虑 nDNA 测序。RNA 测序(转录组学)和呼吸测定法也有助于准确诊断。活检标本通常取自肌肉或皮肤,对于确认血液或尿液样本中可能检测不到的组织特异性 mtDNA 突变仍然很有价值。组织病理学检查和呼吸链酶分析可用于这些组织样本,揭示异常的线粒体结构、形态和功能。因此,活检仍具有重要的诊断价值。
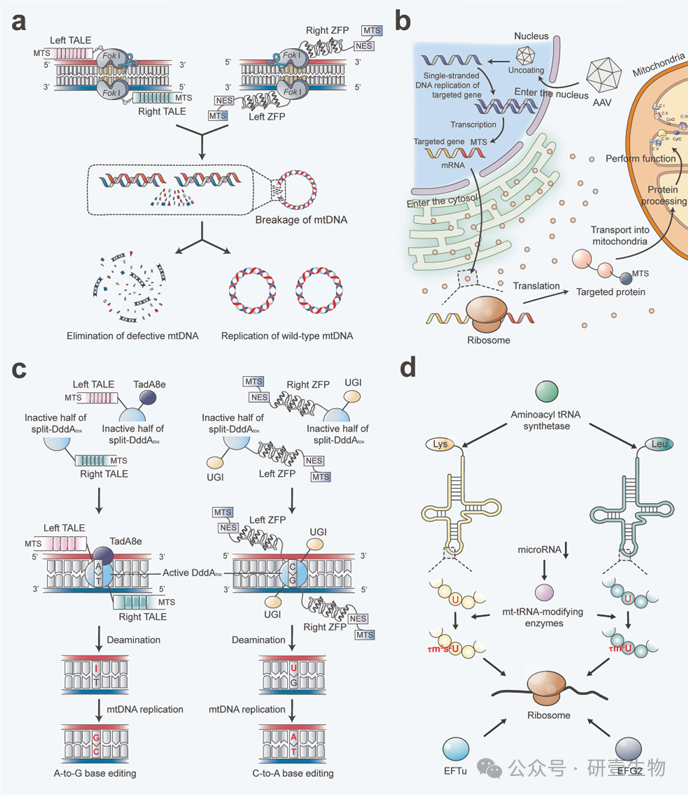
(A) MitoTALENs 由与 FokI 核酸酶融合的 TALE 组成,而 mtZFNs 则由与 FokI 核酸酶连接的 ZFP 组成。MTS 引导 mtZFNs 和 mitoTALENs 进入线粒体。ZFP 和 TALE 选择性地与预先确定的有缺陷的 mtDNA 目标序列结合,然后 FokI 二聚化并切割这些结合位点附近的 mtDNA,造成双链断裂,从而消除有缺陷的 mtDNA。然后,剩余的mtDNA 可以进行复制,从而改变异源比率。 (B) 将目标基因序列与转录调控元件和 MTS 一起打包到 AAV 载体中,然后将其送入受体细胞的细胞核。在细胞核内,AAV 解除包裹并释放出单链 DNA,复制后形成双链 DNA。然后,RNA 聚合酶将 DNA 转录成 mRNA。mRNA 离开细胞核,在细胞质的核糖体上翻译成相应的蛋白质。MTS 引导这些蛋白质进入线粒体,在线粒体中进行进一步加工并发挥各自的功能。(C) DdCBEs 是通过融合 MTS、分裂的 DddAtox 两半、UGIs 以及 ZFPs 或 TALEs 而构建的。DddAtox 催化胞嘧啶脱氨基为尿嘧啶,而 UGIs 则阻止尿嘧啶-DNA 糖基化酶切除尿嘧啶,从而在复制过程中进行 C-T 编辑。此外,通过连接 MTS、分裂-DddAtox 的两半、TadA8e(一种经过改造的腺嘌呤脱氨酶)和 TALEs,TadA8e 可以催化腺嘌呤脱氨成肌苷,而肌苷在复制过程中会与胞嘧啶配对,从而实现有针对性的 A-G 编辑。(D)修饰核编码的氨基酰 tRNA 合成酶的 tRNA 结合域或过表达氨基酰 tRNA 合成酶可提高氨基酰化效率并稳定翻译产物。转录后负调控因子(如 microRNA)的表达可抑制线粒体 RNA 修饰酶的表达,从而影响 mt-tRNA 的修饰。使用 microRNA 拮抗剂有可能逆转疾病表型。此外,过量表达 mt tRNA 修饰酶可以纠正 mt-tRNA 的反密码子第一核苷酸修饰缺陷,从而改善线粒体内核糖体的翻译。线粒体翻译延伸因子 EFTu 和 EFG2 的过表达也能部分抑制翻译延伸过程中 mtDNA 突变导致的氨基酸错结合。
自 1871 年首次描述 LHON 以及随后在 1988 年和 1995 年发现 mtDNA 和 nDNA 在线粒体遗传疾病中的作用以来,该领域已从前分子时代过渡到分子时代。对线粒体生物学的深入了解大大提高了我们阐明发病机制和探索有效疗法的能力。 在了解线粒体疾病的发病机理、改进诊断和开发治疗方法方面取得了重大进展。人们发现了新的线粒体质量控制机制,如迁移体介导的外排、有丝分裂体外排和有丝分裂生成,突出了强大的线粒体质量控制网络在增强线粒体对压力和损伤的恢复能力方面的作用。除了 OXPHOS 干扰外,线粒体功能障碍的广泛影响及其下游效应正受到越来越多的关注。mtDNA或nDNA突变诱发的异常,如有丝分裂-核交流、线粒体动力学和有丝分裂吞噬受损,现已被认为是线粒体疾病的关键因素。值得注意的是,突变特异性和细胞特异性有丝分裂核交流已被证明会显著影响这些疾病中观察到的组织异质性。NGS 和其他技术的进步显著提高了诊断的准确性。前景看好的治疗策略也在不断进步,MRT 在治疗和预防线粒体遗传疾病及其种系传播方面显示出巨大的潜力。ST 技术的成功尤其令人鼓舞。基因治疗,特别是 AAV 介导的异体表达和基因替换,是另一种极具前景的方法,目前有几种疗法正在进行临床试验。 其他干预措施,如运动、药物治疗、细胞疗法、酶替代疗法和器官移植等,都对线粒体疾病产生了有益的影响。然而,一些紧迫的挑战依然存在。线粒体疾病的基因型表型相关性十分复杂,尚未完全明了。 这篇综述系统地总结了线粒体代谢、细胞内和细胞间 MQC 网络、线粒体炎症和凋亡的生理方面。它强调了线粒体疾病的潜在分子机制、最新诊断进展和治疗发展,有助于加深对这些疾病的理解,并指导未来的研究设计和精确诊断与有效治疗的临床转化。本文概述的未来研究、诊断和治疗方向旨在启发和指导后续研究。

